Journal of Psychological Experience
https://doi.org/10.64484/JPsyExp-X4S8M0L2C8N1Y8Z0
Abstract
Keywords: autonomic regulation; Emotional Transformation Therapy; predictive processing; salience network; sensory-salience regulation; thalamocortical coordination
Introduction
A central question in psychotherapy research is how therapeutic interventions produce lasting emotional change. Most explanatory models emphasize cognitive, behavioral, and relational processes, giving less attention to the sensory context in which emotional material is engaged. Yet emotional experience also depends on how the brain prioritizes incoming input, assigns salience, and coordinates information across large-scale networks (Pessoa, 2017; van den Heuvel & Sporns, 2019).
In the present context, salience refers to the prioritization of sensory and internal information across attention, arousal, and action – the assignment of homeostatic relevance to internal and external signals. The nervous system does not merely determine what is emotionally important; it evaluates which bodily sensations and environmental cues matter for regulation, safety, readiness, and adaptive response. Disruptions among the salience network (SN), default mode network (DMN), and executive control network (ECN) have been linked to hypervigilance, rumination, attentional inflexibility, and difficulty shifting between immersive and reflective modes of processing (Menon, 2011; Seeley et al., 2007; Sridharan et al., 2008). From a psychotherapy perspective, therapeutic change may depend not only on shifts in meaning, but on shifts in salience regulation and in the sensory and relational conditions under which experience unfolds.
When distressing emotional material is active, sensory modulation may influence not only what the person notices but also how the organism evaluates the moment in regulatory terms (Barrett & Simmons, 2015; Menon, 2011). Changes in visual stimulation, gaze, brightness, rhythm, or color may help shift an activated state from urgent threat or intolerable arousal toward manageable discomfort or newly tolerable experience (Tamietto & de Gelder, 2010; Vandewalle et al., 2010). The Sensory-Salience Regulation (SSR) therefore proposes that carefully calibrated visual modulation may influence emotional change by shaping early sensory-salience conditions that support emotional engagement, autonomic recalibration, and later reflective integration.
SSR adds a distinct emphasis to existing psychotherapy models. Most accounts explain change through reinterpretation, exposure, skills training, or relational co-regulation. By contrast, SSR highlights the possibility that the sensory conditions of a session may influence the processes that support updating, inhibitory learning, and reconsolidation-consistent change (Craske et al., 2014; Lane et al., 2015). Sensory context warrants explicit consideration because it is continuously present during therapy, closely linked to attentional selection and autonomic state, and open to fine-grained adjustment through light and oculomotor parameters.
Predictive-perception accounts further support the plausibility that structured visual input may influence emotional processing early in the perceptual stream. Human Magnetoencephalography (MEG) studies suggest that prior expectations can induce stimulus-specific sensory templates in visual cortex shortly before an expected stimulus appears, resembling those evoked by actual sensory input (Kok et al., 2017). Visual modulation may therefore bias the sensory-attentional conditions under which emotionally relevant material is first encountered and enters salience processing.
Visual stimulation warrants particular attention because light, together with gaze and oculomotor conditions, provides a sensory medium that can be controlled with unusual precision. Prior research suggests that light can influence arousal, mood, learning, and emotional brain responses (Bedrosian & Nelson, 2017; Fernandez et al., 2018; LeGates et al., 2012; Vandewalle et al., 2010, 2013), while gaze, fixation, and visually guided saccades are linked to attentional selection and thalamic-oculomotor visual processing (McPeek & Keller, 2002; Saalmann et al., 2012; Schneider et al., 2020; Takakuwa et al., 2021). These features make light-based modulation a plausible means of influencing emotional engagement without necessarily disrupting it. Visual stimulation may also remain relevant when its effects are not fully mediated by ordinary conscious seeing, as suggested by evidence from blindsight and visually guided behavior involving residual visual pathways (Ajina & Bridge, 2016; Takakuwa et al., 2021). Broader multisensory research also suggests that visual input interacts with other sensory systems in ways that shape perceptual organization, cognition, and adaptive responding (Murray et al., 2016; Wallace et al., 2020).
Color has been especially difficult to integrate into psychotherapy theory because the physics of light can be measured with precision, whereas the human experience of color cannot. Wavelength and spectral properties are objectively quantifiable, but perceived color is a context-sensitive construction shaped by illumination, contrast, adaptation, surrounding conditions, and observer variability (Foster, 2011; Shevell & Kingdom, 2008). Precisely because color is a perceptual event rather than a fixed property of objects, it may influence salience, affective tone, and the conditions under which emotional material is engaged (Wilms & Oberfeld, 2018).
This article is a narrative review that selectively integrates visual neuroscience, salience-network research, predictive processing, and psychotherapy process theory to examine how sensory modulation may operate during emotional engagement. Its purpose is theory-building rather than systematic evidence synthesis. Emotional Transformation Therapy (ETT) is used as a clinically informative model system for organizing plausible mechanisms and testable predictions, not as evidence that those mechanisms have been established. Because several candidate mechanisms are drawn from animal and basic sensory-neuroscience studies, translational claims are necessarily cautious. Animal findings are treated as mechanistic analogies and sources of hypotheses rather than as direct evidence for ETT mechanisms. Where possible, greater inferential weight is given to human neuroimaging, electrophysiological, psychophysiological, and clinical-process evidence. Within this hypothesis-generating frame, the paper outlines the rationale for sensory-salience regulation, reviews relevant visual and thalamic pathways, examines how visual input may interact with activated emotional states, presents a four-phase SSR framework, and concludes with testable predictions, clinical implications, and future directions.
Emotional Transformation Therapy as a Clinical Model System
ETT, developed by Steven Vazquez, combines conventional psychotherapeutic processing with the systematic variation of visual parameters such as wavelength, brightness, gaze direction, and rhythmic modulation (Vazquez, 2014, 2015). Unlike imagery-based approaches that rely primarily on symbolic or representational content, ETT makes non-representational sensory features explicit and modifiable, creating a distinctive clinical context in which to examine how sensory modulation may interact with emotional processing.
Current Clinical Evidence and Limits of Inference
The ETT-specific evidence base includes substantial clinical experience but remains preliminary in the published research literature. Over nearly two decades, ETT has generated extensive case-based observation among trained therapists, but published reports consist mainly of theoretical descriptions, clinical manuals, case illustrations, and reports of light-based or peripheral eye stimulation procedures (Vazquez, 2005), rather than adequately powered randomized trials, dismantling studies, or independent replications.
This distinction is important for SSR. Clinical reports help identify phenomena that require explanation, including rapid shifts in distress, somatic intensity, perceptual vividness, affective access, and regulatory stability. They do not determine whether such changes reflect ETT-specific visual parameters, therapeutic alliance, expectancy, exposure-like processes, attentional anchoring, therapist allegiance, or other nonspecific factors.
SSR is therefore not presented as evidence that ETT efficacy or mechanisms are established. Rather, ETT is used as a clinically informative model system for generating testable hypotheses. The following vignette illustrates the kinds of perceptual and regulatory shifts reported in ETT clinical work.
In Vazquez's case illustration of "Cara," visual stimulation was adjusted in response to the client's changing emotional state rather than held constant (Vazquez, 2014). As Cara engaged self-critical and attachment-related themes, the viewed colors reportedly became faint, foggy, or disappeared. The therapist treated this shift as a meaningful process cue, briefly redirecting her to the immediate sensory task before emotional exploration resumed. Cara later linked the experience to a longstanding pattern of withdrawal under overwhelming affective conditions: "When others hurt me, I faded".
During treatment, rapid changes in perceived hue, saturation, brightness, or visual clarity may serve as useful indicators of dissociative state shifts (Kusztor et al., 2025; Phelps et al., 2006; Vazquez, 2014). Perceptual flattening, muting, or fading may suggest movement toward dissociative disengagement, whereas the return of greater vividness and differentiation may indicate renewed embodied presence and emotional access (Barrett & Simmons, 2015). These observations are best understood as hypothesis-generating process markers rather than as disorder-specific indicators.
From the perspective of SSR, such shifts suggest that visual experience may change along with the client's emotional and regulatory state. Targeted visual input may therefore function as a form of attentional anchoring, helping emotional material remain organized, tolerable, and therapeutically accessible while reducing the likelihood of flooding or defensive withdrawal. ETT thus provides a concrete clinical setting in which sensory modulation is co-regulated through therapist attunement and emotional processing.
Three features of ETT are especially relevant. First, visual input may influence emotional processing through preconscious pathways and need not depend entirely on ordinary conscious seeing. Second, when distressing material is active, modulated visual input may become incorporated into the unfolding emotional state rather than merely accompany it. Third, ETT uses not only the titration of light parameters but also gaze direction, fixation, and guided eye movement to shape attention and salience.
These ETT-specific procedures can be mapped only provisionally onto the SSR phases. Peripheral Eye Stimulation (PES) varies peripheral light input and eye position while the client remains engaged with affective, cognitive, or somatic material, with the clinical aim of reducing flooding, dissociation, or somatic distress (Vazquez, 2005); within SSR, PES is most relevant to Phases 1 through 3 because it may influence early sensory priority, modify the active emotional field, and support shifts in bodily salience and regulation. Multidimensional Eye Movement (MDEM) combines fixed and guided eye positions with wavelength variation, three-dimensional focal points, and therapist responsiveness during activated material (Vazquez, 2009); within SSR, MDEM is most relevant to Phases 1, 2, and 4. These mappings are procedure-specific hypotheses rather than established mechanisms, and clarifying how PES, MDEM, and other ETT procedures differentially engage the proposed SSR phases remains a priority for future research.
This provisional mapping leads to the broader SSR framework. Within SSR, sensory refers to incoming signals from the external environment and the body, salience to the prioritization of those signals as relevant within an activated context, and regulation to the dynamic adjustment of emotional, physiological, and attentional responding. The four overlapping phases are heuristic rather than fixed neural stages, presented sequentially for clarity but understood as dynamically interacting within an attuned therapeutic context.
In Phase 1, targeted visual input enters the same experiential field as emotional content and influences early sensory prioritization. In Phase 2, managed sensory mismatch alters the weighting of current input against rigid threat-based expectations while emotional engagement is maintained. In Phase 3, the active pattern shifts toward a more regulated bodily trajectory, linking salience processing with autonomic and interoceptive regulation. In Phase 4, large-scale coordination supports reflective distance, autobiographical reorganization, and adaptive updating. Figure 1 provides a conceptual summary.
Figure 1
Four Phases of Sensory-Salience Regulation in Emotional Transformation Therapy
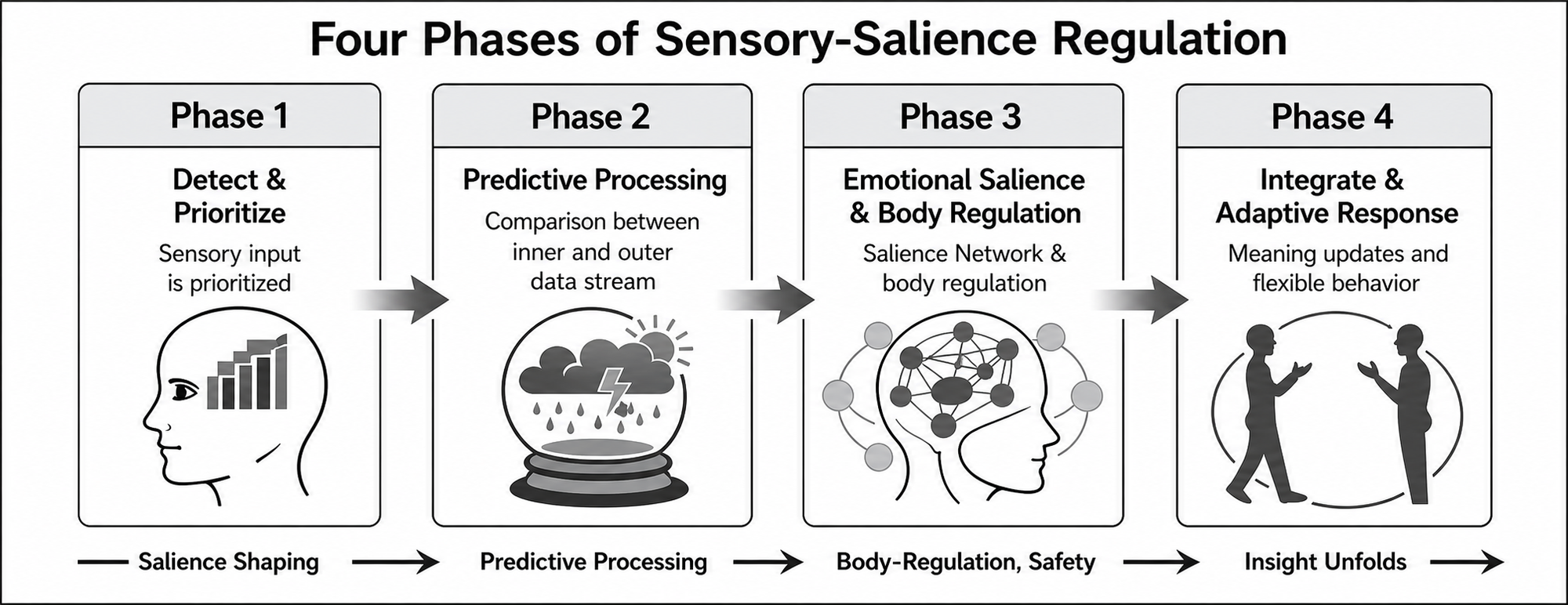
Note. Within therapist attunement, co-regulation, titrated visual modulation, and the client's regulatory window, activated distress is proposed to move through four partially overlapping phases: (1) salience shaping, in which sensory input is detected and prioritized; (2) predictive processing, in which internal and external streams of information are compared and the expected emotional trajectory begins to shift; (3) body recoding and safety, in which body-emotional regulation and salience-network coordination become more stable; and (4) insight and updating, in which adaptive meaning-making, reflective access, and flexible responding become more possible. These processes may be reflected in reduced reactivity, greater affect tolerance, improved reflective access, and more adaptive functioning.
Phase 1: Co-activation and Early Salience Shaping
The first phase of the SSR model begins once emotional material is active and targeted visual input enters the same moment-to-moment field of experience. The central claim is that visual stimulation is processed in relation to the ongoing emotional state rather than as incidental background. Once a painful memory, self-state, or body-state pattern is engaged in session, changes in visual conditions become part of the field in which that material is experienced and observed.
Recent work supports the plausibility of such interaction. Internally generated imagery and low-strength external visual input can combine within shared perceptual systems to shape a graded perceptual-reality signal influencing whether experience feels more or less real in the present moment; Dijkstra et al. (2025) linked this effect to bilateral fusiform cortex. Within SSR, this suggests that non-representational visual modulation may influence the vividness, immediacy, and accessibility of activated material rather than merely serving as nonspecific background.
Phase 1 therefore concerns how visual input begins to shape attentional priority within an already activated field. Research on visual salience suggests that local feature contrast can increase attentional pull, with early physiological responses and cortical (V1) activity tracking the strength of that effect (Zhang et al., 2012). Structured visual stimulation may thus influence which aspects of the active field are most likely to capture attention and organize experience during emotional activation.
Color may contribute to early salience shaping because hue, brightness, and saturation influence attentional deployment and arousal, and because human color processing extends through ventral occipitotemporal cortex, including fusiform-related territory (Beauchamp et al., 1999; McKeefry & Zeki, 1997). Subsequent intracranial, stimulation, and fMRI findings support the involvement of anterior and ventral color-biased regions when color becomes perceptually or behaviorally relevant (Lafer-Sousa et al., 2016; Murphey et al., 2008). This does not imply that particular colors correspond to fixed emotional meanings; rather, color modulation may enter perceptual systems that shape how active material is represented, captures attention, and feels vivid or immediate.
This interpretation is consistent with evidence that color vividness can enhance early indices of attentional deployment, while chromatic- and luminance-based salience make partly distinct contributions to visual priority (Forschack et al., 2025; Hardman et al., 2020). Saturated and brighter colors appear more likely to increase arousal, with hue contributing additional variation in emotional tone (Wilms & Oberfeld, 2018), and color-emotion correspondences reflect the combined influence of lightness, saturation, and hue rather than hue alone (Jonauskaite & Mohr, 2025). SSR, therefore, treats color effects as context-sensitive and multidimensional rather than as evidence for a universal symbolic color code.
Color may also influence perceived immediacy: brighter and more saturated inputs tend to support relative nearness, whereas reduced contrast and saturation are associated with greater apparent distance (Dresp-Langley & Reeves, 2014), and visual salience and perceived depth jointly influence attentional priority (Qian et al., 2018). Recent optics work further suggests that wavelength composition can influence the accommodative selection of which information is brought into best focus (Chin et al., 2026), making it plausible that color modulation affects the immediacy or depth-like quality of experience at an early sensory level, before semantic interpretation.
Several converging mechanisms may support this early prioritization before fuller appraisal is complete. Contemporary accounts suggest that emotionally relevant input can bias orienting and bodily readiness quickly while other pathways support contextually informed perception, making it more useful to think in terms of interacting routes than of a rigid low-road/high-road split (Pessoa & Adolphs, 2010). Structured visual stimulation may therefore alter salience before the emotional pattern itself has been revised.
Evidence from affective neuroscience suggests that emotionally relevant visual signals can influence orienting and salience processing before full semantic appraisal is complete (McFadyen et al., 2017; Tamietto & de Gelder, 2010). One likely contributor is the superior colliculus, a midbrain system involved in rapid orienting toward potentially important events: human imaging shows increased activity during aversive relative to neutral visual processing (Wang et al., 2020), and animal and nonhuman primate studies indicate feature-sensitive, graded responses to contrast, temporal frequency, and color-related change (Calanni et al., 2024; Chen & Hafed, 2018; Herman & Krauzlis, 2017). Related work also indicates that superior colliculus circuitry can rapidly engage autonomic adjustments associated with orienting, linking early visual salience to bodily readiness (Lynch et al., 2022). These findings do not establish that superior colliculus mechanisms explain psychotherapeutic change; they identify feature-sensitive orienting systems whose dimensions overlap with stimulus features manipulated in ETT.
Thalamic systems may also help shape this phase. Visual processing depends not only on cortical analysis but also on thalamic mechanisms that contribute to attentional selection and the coordination of communication across cortical areas (Halassa & Kastner, 2017; Sherman, 2016). The lateral geniculate nucleus may help visual information continue to be processed even when ordinary conscious vision is reduced (Schmid et al., 2010), while the pulvinar has been implicated in attentional selection and distractor filtering and may help regulate information flow across cortical systems (Strumpf et al., 2013). Multimodal findings showing an early transient pulse of activity in midbrain reticular formation and central thalamus across sensory modalities further support the possibility that modulated sensory input can shape the conditions under which attention and emotional processing unfold (Khalaf et al., 2025).
This is especially relevant to ETT because visual stimulus dimensions that can be directly manipulated – brightness, saturation, flicker, and spatial position relative to gaze – fall within the range of features capable of influencing early orienting and attentional systems. In practice, these parameters may help determine whether activated material becomes overwhelming or remains manageable enough to engage without escalating distress.
Phase 1 thus concerns priority setting rather than reinterpretation: what stands out, what feels immediate, and what remains tolerable enough to stay in awareness. Phase 2 begins once these altered conditions of priority start to influence the expected course of the active pattern.
Phase 2: Precision Reweighting and Mismatch
Whereas Phase 1 concerns access and early salience shaping, Phase 2 concerns what happens once that now-prioritized input begins to alter the expected course of the active emotional state. Once a memory, belief, or body-state pattern is activated, the system is not simply replaying the past; it is also generating expectations about what will happen next – for example, that danger will escalate, distress will become unmanageable, or the person will again be pulled into a familiar maladaptive state. Emotional schemas can, therefore, be understood as organized predictive patterns shaped by prior learning.
Perception is continuously shaped by the interaction between incoming signals and prior expectations. Predictive-processing accounts describe this as an ongoing comparison between top-down predictions and bottom-up input, with mismatch signals updating the system when current evidence diverges from what was expected (Clark, 2013; Friston, 2010; Keller & Mrsic-Flogel, 2018). Change becomes possible when sensory input is similar enough to keep the emotional pattern engaged, yet different enough to alter its expected course. The central process in this phase is therefore managed sensory mismatch: a tolerable difference introduced into an otherwise rigid emotional sequence. The client remains in contact with painful material, but the experience no longer unfolds in quite the same way. Its distinctive contribution is not simply a change in activation, but a loosening of the expectation that the emotional sequence must continue along its familiar course.
In predictive-processing terms, this corresponds to a shift in precision weighting, in which confidence begins to move away from prior expectations and toward current sensory evidence (Kanai et al., 2015). Visual mismatch studies show that unexpected changes in simple sensory features can generate prediction-error responses rather than merely generalized reactivity (Stefanics et al., 2018). Recent thalamocortical work found that unexpected visual input can selectively amplify unpredicted sensory features, with this amplification depending on cooperative pulvinar input and local inhibitory gating (Furutachi et al., 2024). Higher-order thalamic systems may help regulate the gain assigned to prediction and prediction-error signals, thereby influencing which sensory cues are treated as informative enough to guide perception and behavior (Kanai et al., 2015).
Recent animal findings extend this logic in a treatment-relevant direction. In a mouse model, a single low-contrast visual threat cue later produced marked avoidance of a previously neutral location, despite producing no measurable freezing during the original exposure. This later avoidance required visual input at testing and depended on melanopsin-sensitive signaling through a light-responsive pathway linked to contextual avoidance rather than immediate defensive responding (Aranda et al., 2026). Related human findings support the broader translational logic: visually specified contexts can acquire threat value and later guide avoidance behavior, and fear learning can alter visuocortical processing of contextual cues themselves (Glotzbach et al., 2012; Santos-Mayo & Moratti, 2025).
Clinically, the significance of Phase 2 is that the emotional pattern begins to lose some of its predictive grip while engagement is maintained. In ETT, visual modulation may introduce small but meaningful deviations into an otherwise rigid emotional-perceptual loop. The client remains within the emotional material, but it no longer unfolds under exactly the same predictive rules. This helps explain how therapeutic change may begin before the person arrives at an explicit cognitive reinterpretation (Elsey et al., 2018; Lane et al., 2015). What may begin shifting first is not the explicit meaning of the experience, but its felt trajectory: what seems likely to happen next, how compelling avoidance feels, and how strongly the state organizes perception, attention, and action.
Perceptual changes such as dimming or reduced vividness may serve as process markers that precision weighting is shifting while emotional material remains active, consistent with evidence that affective arousal can alter early visual processing and contrast sensitivity, and that derealization is often accompanied by fogginess and altered color intensity (Kusztor et al., 2025; Phelps et al., 2006). The therapist's task is to help the client remain in workable contact with the emerging experience so that mismatch remains informative rather than overwhelming. Phase 3 begins once this shift starts to register more directly in the body.
Phase 3: Body Re-coding and Salience Regulation
Whereas Phase 2 concerns the destabilization of rigid threat-based predictions through calibrated sensory mismatch, Phase 3 describes how those shifts begin to register more directly in the body. Here, the dominant level of change moves from predictive inference toward embodied regulation, as the emotional pattern begins to anticipate a more regulated physiological trajectory. Many maladaptive schemas are maintained not only by remembered meanings but also by recurrent body-state predictions such as tightening, dread, autonomic escalation, collapse, or defensive withdrawal. Contemporary models of interoception suggest that such anticipated bodily states are integral to emotional experience; distress may be sustained as much by an expected bodily future as by a remembered past (Barrett & Simmons, 2015; Paulus & Stein, 2010).
In the SSR model, body re-coding refers to a shift in the expected bodily trajectory toward greater autonomic and interoceptive stability while emotional material remains engaged (Thayer & Lane, 2000). This shift is proposed to involve not only changes in arousal but also changes in salience-network coordination and body-state representation. In a safe and attuned context, the client does not merely think differently about a memory; the memory begins to unfold toward a more regulated physiological future, with less defensive narrowing and greater affect tolerance.
As the activated state becomes less rigidly organized, structured visual input may begin to alter not only arousal but also the weighting and integration of sensory signals that contribute to bodily self-experience. Visuo-proprioceptive research suggests that visual input can bias perceived body position and support short-term recalibration under varying contextual conditions (Kirsch & Kunde, 2023; Wali et al., 2023). This is especially relevant to ETT, where visual stimulation, gaze direction, and eye position are often used to engage specific body zones linked to pain or emotional distress, sometimes with rapid shifts in bodily intensity, localization, or affective charge – changes that may reflect not only altered arousal but a reorganization of bodily salience.
Phase 3 also marks a shift from salience amplification toward a closer coupling between salience and regulation. What feels urgent or threatening no longer escalates automatically, and the client becomes better able to remain with the experience without being overwhelmed by it. A likely contributor is the salience network – especially the insula and dorsal anterior cingulate, which help register what is most important and coordinate shifts in attentional and regulatory state (Menon & Uddin, 2010; Seeley et al., 2007). Posterior insular and nearby multisensory regions may help integrate signals related to bodily position, orientation, and self-motion (Cai & Menon, 2025; Liu & Zeng, 2025), while more anterior insular regions and related salience-network structures may translate those bodily changes into altered feelings of importance, attentional focus, and a broader shift toward a more regulated mode of engagement. Phase 3 may therefore reflect not just reduced arousal, but a change in how bodily experience is registered, prioritized, and managed.
Neurovisceral integration models are consistent with this account, proposing that attentional and autonomic regulation are tightly linked, with more adaptive functioning associated with greater physiological flexibility (Thayer & Lane, 2000). Human imaging suggests that autonomic responses to emotionally salient stimuli involve coordinated activity across the amygdala, insula, anterior cingulate, and brainstem (Critchley et al., 2005). More recent work suggests that during negative emotional processing, some systems intensify the felt urgency of activated material, whereas others help prevent that activation from escalating into unchecked bodily arousal (Ince et al., 2023). In mood and anxiety disorders, this regulatory containment appears reduced (Ince et al., 2025), consistent with the possibility that Phase 3 reflects a shift away from unchecked salience escalation toward a more regulated coupling of bodily arousal, attentional priority, and emotional experience.
Phase 3 therefore concerns changing the bodily organization of emotional salience rather than suppressing emotion. The client remains in contact with the experience yet is less likely to be drawn into the familiar bodily trajectory of the activated pattern; distress may still be present, but is less likely to cascade into flooding, collapse, or dissociative disengagement. Therapist attunement is therefore not secondary to sensory intervention but one of the conditions that makes this phase workable. Research on interpersonal synchrony suggests that therapists and clients can co-regulate across physiological channels (Bar-Kalifa et al., 2023; Koole & Tschacher, 2016). ETT can thus be understood as a co-regulated sensory intervention in which visual modulation helps shift salience and perceptual weighting, while the therapeutic relationship helps those changes be experienced as safe and adaptively engaging. As bodily expectancy becomes less rigid and salience becomes more tightly coupled with regulation, the conditions emerge for broader changes in meaning and self-organization.
Phase 4: Integration, Insight, and Updating
Phase 4 describes the emergence of broader integrative processing once salience and bodily regulation have become more stable. In this phase, organization shifts toward large-scale network coordination, supporting reflective distance, autobiographical reorganization, and meaning-based updating (Andrews-Hanna et al., 2014; Menon, 2011; Sridharan et al., 2008). Earlier changes in sensory prioritization, predictive mismatch, and bodily regulation now support a more flexible and integrated relationship to emotional experience as a whole. Once a person is affectively engaged but no longer dominated by raw reactivity, previously distressing material may begin to be encountered differently – not simply as something to endure, but as something that can be observed, reflected upon, and reorganized. Clients may report not only feeling calmer but also sensing a broader autobiographical perspective and a changed relationship to what had previously felt overwhelming. Insight is, therefore, understood not as a separate cognitive event imposed from above, but as an emergent expression of successful sensory-affective reorganization (Lane et al., 2015).
A large-scale network perspective helps frame this phase. Menon's Triple-Network Model proposes that the salience network detects what is most relevant, the executive control network supports reflective regulation, and the default mode network contributes to autobiographical meaning and self-integration (Andrews-Hanna, 2014; Menon, 2011). In rigid emotional states, movement between internal absorption and present-moment engagement may become inflexible, leaving the person either submerged in distress or cut off from it through defensive constriction. Therapeutic change therefore depends in part on restoring more flexible, salience-guided movement across these modes.
The right anterior insula and mediodorsal thalamus have been implicated in such transitions, helping bias the brain toward externally directed engagement or internally focused cognition (Bolton et al., 2020; Harrison et al., 2022), and mediodorsal thalamic and ventral pallidal systems may contribute to subcortical regulation of DMN-related states (Zhao et al., 2024). As salience becomes less dominated by threat, reflective capacities that were previously narrowed, inaccessible, or quickly overwhelmed may become more available.
Autobiographical memory may therefore be reorganized with less fusion and defensive constriction. Cognitive and autobiographical change do not stand apart from earlier phases but emerge from the ongoing reorganization of sensory, salience, and regulatory processes. The result is not simply reduced activation, but a greater capacity to locate distressing material within a broader sense of self, time, and meaning.
The SSR model does not propose a fixed neural sequence. Rather, earlier sensory conditions are proposed to bias salience, thalamic filtering to shape that bias, and salience-network dynamics to influence broader network switching (Cortes et al., 2026), creating the conditions under which reflective processing and a more adaptive sense of self can emerge.
Testable Predictions
If the SSR framework is broadly correct, targeted sensory modulation introduced during affectively engaged psychotherapy should produce measurable differences across neural, psychophysiological, and behavioral domains.
Phase 1 – Co-activation and Salience Shaping
Candidate indices include lateralized alpha activity, attentional ERP markers such as the N2-posterior-contralateral (N2pc), and overt attentional allocation measured through eye tracking, with neuroimaging used where appropriate to examine related visual-attentional circuitry.
Phase 2 – Precision Reweighting and Mismatch
Candidate indices include ERP mismatch responses, pupillometry, expectancy ratings, and behavioral markers of sustained engagement such as gaze persistence or reduced avoidance.
Phase 3 – Body Re-coding and Salience Regulation
Predicted indices include coordinated changes in anterior insula and dorsal anterior cingulate cortex (dACC) recruitment together with psychophysiological measures of autonomic regulation such as heart-rate variability.
Phase 4 – Integration, Insight, and Updating
Broader network-level changes are predicted, including more flexible SN-DMN-ECN coordination and behavioral indicators such as reduced reactivity, greater decentering or autobiographical perspective, and improved functioning.
Table 1 summarizes these phase-specific predictions, linking each phase to candidate systems, methods, and expected patterns of change.
Table 1
Phase-Specific Predictions Derived from the Sensory-Salience Regulation Framework
| Model phase | Candidate systems / processes | Method | Predicted change |
|---|---|---|---|
| Co-activation and salience shaping | LGN, pulvinar, superior colliculus pathways, thalamocortical gain | ERP, EEG/MEG, eye tracking, fMRI | Altered early sensory prioritization and salience allocation |
| Precision reweighting and mismatch | Pulvinar, predictive processing, mismatch signaling | ERP, pupillometry, expectancy ratings, engagement measures | Reduced threat certainty, sustained engagement, increased awareness |
| Body re-coding and salience regulation | Anterior insula, dACC, salience network, neurovisceral integration | Task-based fMRI, HRV, psychophysiology | Reduced hyperreactivity, greater autonomic flexibility |
| Integration, insight, and updating | SN-DMN-ECN coordination and network flexibility | Resting-state fMRI, connectivity analysis, clinical follow-up | More flexible network coordination, reduced reactivity, improved functioning |
Note. dACC = dorsal anterior cingulate cortex; DMN = default mode network; ECN = executive control network; EEG = electroencephalography; ERP = event-related potential; fMRI = functional magnetic resonance imaging; HRV = heart-rate variability; LGN = lateral geniculate nucleus; MEG = magnetoencephalography; SN = salience network.
Discussion
The present review examined how sensory modulation may shape emotional processing when distressing material is actively engaged in psychotherapy. The Sensory-Salience Regulation (SSR) framework was developed as a hypothesis-generating model that links visual input, salience regulation, predictive processing, bodily regulation, and later reflective integration. The following discussion summarizes the clinical, theoretical, and research implications of this framework while emphasizing that the proposed mechanisms remain provisional and require direct empirical testing.
The SSR framework suggests that sensory modulation can help sustain engagement with affective experience long enough for reflective processing and updating to occur (Lane et al., 2015; Thayer & Lane, 2000). Within SSR, sensory modulation is understood as an explicit component of the therapeutic process rather than as a separate mechanism of change. Visual stimulation, gaze, fixation, and guided eye movement may shape how emotional material is accessed, weighted, and regulated in real time through therapist attunement, co-regulation, and attention to the client's window of tolerance (Bar-Kalifa et al., 2023; Koole & Tschacher, 2016).
Observing perceptual, interoceptive, and bodily shifts during therapy may provide useful cues for pacing and formulation. Changes in brightness, color clarity, bodily tension, or autonomic settling may help indicate whether emotionally salient material is becoming more tolerable, more dysregulated, or more available for integration.
More broadly, visual stimulation can remain relevant to treatment even when its effects are not fully mediated by ordinary conscious seeing (Ajina & Bridge, 2016; Takakuwa et al., 2021). Light may influence orienting, arousal, and attentional selection through preconscious visual and subcortical-thalamic pathways (Fernandez et al., 2018; Vandewalle et al., 2013), and once distressing material is active, such modulation may become incorporated into the unfolding emotional state itself, influencing salience, conscious access, and the circumstances under which that state is re-entered and potentially revised (Dijkstra et al., 2025; Murray et al., 2016; Wallace et al., 2020).
Gaze direction, prolonged fixation, and guided eye movement may extend this regulatory range further by shaping how sensory information is prioritized and maintained within the emotional field (McPeek & Keller, 2002; Saalmann et al., 2012; Schneider et al., 2020). Related methods such as Eye Movement Desensitization and Reprocessing (EMDR) and Brainspotting suggest that gaze-related procedures can influence emotional processing, while ETT-specific procedures such as PES and MDEM point to a broader use of gaze-dependent modulation (Vazquez, 2005).
Within a supportive therapeutic setting, shifts in visual parameters such as hue, saturation, brightness, rhythm, and eye-gaze position may help alter the early salience conditions under which emotionally charged material is re-entered. Rather than directly "erasing" fear, these adjustments may help keep the client engaged with previously threatening material at a tolerable level, allowing mismatch between anticipated danger and present-moment safety to be registered, supporting contextual safety learning and reducing the immediate threat value of older emotional memories.
On this view, clinically observed color shifts in ETT need not reflect a symbolic color code but may instead arise from dynamic changes in color-biased perceptual systems when color becomes behaviorally or emotionally relevant. The proposed direction of change is toward greater bodily regulation, less defensive escalation, and more flexible engagement with emotionally salient material.
SSR is also generally consistent with memory reconsolidation, inhibitory-learning, predictive-processing, and neurovisceral integration models, while making the sensory context of emotional engagement more mechanistically explicit. Higher-order thalamic systems, particularly the pulvinar, may help regulate how competing sensory signals are weighted as an emotional pattern is re-entered and revised, with broader transthalamic routing helping coordinate attentional selection and information flow across cortical systems.
The model is also bidirectional. Prefrontal and cortical influences on subcortical defensive responding suggest that meaning-making and relational co-regulation may interact with sensory modulation even during early phases of salience shaping. SSR, thus, extends psychotherapy models centered on cognitive, behavioral, and relational processes by proposing that early sensory processing, attentional weighting, and salience regulation may also contribute directly to therapeutic change.
Clinically, this synthesis suggests that perceptual, interoceptive, and autonomic shifts may serve as useful process cues for titrating emotional engagement within an attuned therapeutic relationship. It also frames visual and gaze-related modulation as potentially useful only when embedded in careful pacing, co-regulation, and attention to the client’s regulatory capacity. Theoretically, SSR links sensory context, salience regulation, predictive processing, neurovisceral regulation, and reflective integration within a unified model of emotional change. Empirically, it points to studies testing whether specific visual, gaze, and rhythmic parameters alter salience, autonomic regulation, and reflective integration during emotionally engaged psychotherapy. Future studies should use active comparison conditions and multimodal measures to distinguish SSR-specific predictions from common therapeutic factors.
Limitations and Boundary Conditions
Several limitations and boundary conditions should be noted. First, the proposed sequence is theoretical and has not been directly established in ETT through controlled trials. Second, the phase-organizing language is intended as a clinical heuristic rather than as a claim that the visual-salience literature supports one fixed model of salience processing. Third, structures such as the pulvinar are best understood within interacting systems rather than as isolated centers of change. Fourth, the framework does not assume that any specific wavelength or visual parameter has a universal effect; individual differences in sensory processing, trauma history, and regulatory capacity are likely to shape response.
As a narrative review, this article is vulnerable to confirmation bias in evidence selection and does not rule out null findings, contradictory evidence, or alternative explanations. Apparent ETT-associated improvements could reflect therapeutic alliance, expectancy, emotional exposure, demand characteristics, or the stabilizing effects of an absorbing sensory task rather than the specific effects of visual modulation. Common factors therefore remain an alternative explanation for psychotherapy outcomes (Wampold, 2015; Cuijpers et al., 2019). Evidence that eye movements may contribute to emotional-memory processing in EMDR does not establish that any specific eye position, movement pattern, wavelength, or light parameter is necessary or sufficient for change in ETT (Lee & Cuijpers, 2013). Future studies should use active comparison conditions matching therapist contact, expectancy, emotional activation, visual engagement, and attentional demand while varying the proposed sensory parameters.
A further limitation concerns translational inference. Some evidence supporting Phases 1 and 2 comes from animal or nonhuman primate studies of visual orienting, superior colliculus function, pulvinar-mediated attention, and sensory prediction error. These help identify basic mechanisms that are difficult to isolate during psychotherapy but cannot determine how such processes unfold in human treatment. Future studies should prioritize human EEG/MEG, eye tracking, pupillometry, HRV, and neuroimaging during emotionally engaged sensory modulation.
The role of color in psychotherapy also remains difficult to formalize. The color literature in psychological functioning is less developed than the literature on color physics, physiology, and neuroscience, and color effects are easily confounded unless hue, lightness, and saturation are carefully distinguished. Within SSR, responses to visual modulation are expected to vary across individuals, with temperament, sensory sensitivity, autonomic state, and treatment context shaping whether a given parameter is experienced as activating, tolerable, or regulating. Current evidence supports subcortical and cortical sensitivity to chromatic and hue-relevant information but does not support universal emotional effects of specific wavelengths or hues. Any account of specific colors within SSR should therefore remain provisional, context-sensitive, and empirically testable.
Finally, sensory mismatch alone does not guarantee reconsolidation or therapeutic updating. For the proposed sequence to unfold therapeutically, the intervention must remain within the client's window of tolerance and be supported by therapist attunement, timing, and titration.
Conclusion
The Sensory-Salience Regulation (SSR) framework offers a clinically grounded, neurobiologically informed account of how targeted sensory modulation may influence emotional processing in psychotherapy. It proposes that non-representational visual stimulation may alter the context in which distressing material is re-engaged, regulated, and potentially revised, linking moment-to-moment clinical phenomena with plausible underlying mechanisms. The four-phase model further clarifies this process. When emotional material is active, modulated sensory stimulation may alter access and salience, loosen rigid predictive organization through manageable mismatch, support a more regulated bodily trajectory, and enable broader integration, insight, and updating. Under these conditions, sensory modulation may help sustain engagement with distressing material while reducing the likelihood of overwhelming defensive escalation. Although the framework remains investigational, it organizes a range of observed effects into a coherent and testable model. If supported by future research, SSR could broaden contemporary psychotherapy theory by suggesting that sensory-salience regulation is a meaningful pathway of therapeutic change.
References
Ajina, S., & Bridge, H. (2016). Blindsight and unconscious vision: what they teach us about the human visual system. The Neuroscientist, 23(5), 529–541. https://doi.org/10.1177/1073858416673817
Andrews‐Hanna, J. R., Smallwood, J., & Spreng, R. N. (2014). The default network and self‐generated thought: component processes, dynamic control, and clinical relevance. Annals of the New York Academy of Sciences, 1316(1), 29–52. https://doi.org/10.1111/nyas.12360
Aranda, M. L., Min, E., Liu, L. T., Schipma, A. E., Wang, H., & Schmidt, T. M. (2026). Light tunes long-term threat avoidance behavior in male mice. Nature Communications, 17(1). https://doi.org/10.1038/s41467-026-69564-0
Bar-Kalifa, E., Goren, O., Gilboa-Schechtman, E., Wolff, M., Rafael, D., Heimann, S., Yehezkel, I., Scheniuk, A., Ruth, F., & Atzil-Slonim, D. (2023). Clients’ emotional experience as a dynamic context for client–therapist physiological synchrony. Journal of Consulting and Clinical Psychology, 91(6), 367–380. https://doi.org/10.1037/ccp0000811
Barrett, L. F., & Simmons, W. K. (2015). Interoceptive predictions in the brain. Nature Reviews Neuroscience, 16(7), 419–429. https://doi.org/10.1038/nrn3950
Beauchamp, M. S., Haxby, J. V., Jennings, J. E., & DeYoe, E. A. (1999). An fMRI version of the Farnsworth–Munsell 100-Hue test reveals multiple color-selective areas in human ventral occipitotemporal cortex. Cerebral Cortex, 9(3), 257–263. https://doi.org/10.1093/cercor/9.3.257
Bedrosian, T. A., & Nelson, R. J. (2017). Timing of light exposure affects mood and brain circuits. Translational Psychiatry, 7(1), e1017–e1017. https://doi.org/10.1038/tp.2016.262
Bolton, T. A. W., Wotruba, D., Buechler, R., Theodoridou, A., Michels, L., Kollias, S., Rössler, W., Heekeren, K., & Van De Ville, D. (2020). Triple Network Model Dynamically Revisited: Lower salience network state switching in pre-psychosis. Frontiers in Physiology, 11. https://doi.org/10.3389/fphys.2020.00066
Cai, W., & Menon, V. (2025). Heterogeneity of human insular cortex: five principles of functional organization across multiple cognitive domains. Brain Structure and Function, 230(8). https://doi.org/10.1007/s00429-025-03027-y
Calanni, J. S., Aranda, M. L., Dieguez, H. H., Dorfman, D., Schmidt, T. M., & Rosenstein, R. E. (2024). An ethologically relevant paradigm to assess defensive response to looming visual contrast stimuli. Scientific Reports, 14(1). https://doi.org/10.1038/s41598-024-63458-1
Chen, C.-Y., & Hafed, Z. M. (2018). Orientation and Contrast Tuning Properties and Temporal Flicker Fusion Characteristics of Primate Superior Colliculus Neurons. Frontiers in Neural Circuits, 12. https://doi.org/10.3389/fncir.2018.00058
Chin, B. M., Banks, M. S., Nankivil, D., Roorda, A., & Cooper, E. A. (2026). Focusing on color: How the eye chooses which wavelength to see best. Science Advances, 12(14). https://doi.org/10.1126/sciadv.aea5693
Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behavioral and Brain Sciences, 36(3), 181–204. https://doi.org/10.1017/s0140525x12000477
Cortes, N., Mandel, C., Ikan, L., & Casanova, C. (2026). The pulvinar nucleus and its role in cognitive functions. Brain Research Bulletin, 234, 111672. https://doi.org/10.1016/j.brainresbull.2025.111672
Craske, M. G., Treanor, M., Conway, C. C., Zbozinek, T., & Vervliet, B. (2014). Maximizing exposure therapy: An inhibitory learning approach. Behaviour Research and Therapy, 58, 10–23. https://doi.org/10.1016/j.brat.2014.04.006
Critchley, H. D., Rotshtein, P., Nagai, Y., O’Doherty, J., Mathias, C. J., & Dolan, R. J. (2005). Activity in the human brain predicting differential heart rate responses to emotional facial expressions. NeuroImage, 24(3), 751–762. https://doi.org/10.1016/j.neuroimage.2004.10.013
Cuijpers, P., Reijnders, M., & Huibers, M. J. H. (2019). The Role of Common Factors in Psychotherapy Outcomes. Annual Review of Clinical Psychology, 15(1), 207–231. https://doi.org/10.1146/annurev-clinpsy-050718-095424
Dijkstra, N., von Rein, T., Kok, P., & Fleming, S. M. (2025). A neural basis for distinguishing imagination from reality. Neuron, 113(15), 2536–2542. https://doi.org/10.1016/j.neuron.2025.05.015
Dresp-Langley, B., & Reeves, A. (2014). Effects of saturation and contrast polarity on the figure-ground organization of color on gray. Frontiers in Psychology, 5. https://doi.org/10.3389/fpsyg.2014.01136
Elsey, J. W. B., Van Ast, V. A., & Kindt, M. (2018). Human memory reconsolidation: A guiding framework and critical review of the evidence. Psychological Bulletin, 144(8), 797–848. https://doi.org/10.1037/bul0000152
Fernandez, D. C., Fogerson, P. M., Lazzerini Ospri, L., Thomsen, M. B., Layne, R. M., Severin, D., Zhan, J., Singer, J. H., Kirkwood, A., Zhao, H., Berson, D. M., & Hattar, S. (2018). Light Affects Mood and Learning through Distinct Retina-Brain Pathways. Cell, 175(1), 71–84. https://doi.org/10.1016/j.cell.2018.08.004
Forschack, N., Oxner, M., & Müller, M. M. (2025). The consequences of color chromaticity on electrophysiological measures of attentional deployment in visual search. iScience, 28(4), 112252. https://doi.org/10.1016/j.isci.2025.112252
Foster, D. H. (2011). Color constancy. Vision Research, 51(7), 674–700. https://doi.org/10.1016/j.visres.2010.09.006
Friston, K. (2010). The free-energy principle: a unified brain theory? Nature Reviews Neuroscience, 11(2), 127–138. https://doi.org/10.1038/nrn2787
Furutachi, S., Franklin, A. D., Aldea, A. M., Mrsic-Flogel, T. D., & Hofer, S. B. (2024). Cooperative thalamocortical circuit mechanism for sensory prediction errors. Nature, 633(8029), 398–406. https://doi.org/10.1038/s41586-024-07851-w
Glotzbach, E., Ewald, H., Andreatta, M., Pauli, P., & Mühlberger, A. (2012). Contextual fear conditioning predicts subsequent avoidance behaviour in a virtual reality environment. Cognition & Emotion, 26(7), 1256–1272. https://doi.org/10.1080/02699931.2012.656581
Halassa, M. M., & Kastner, S. (2017). Thalamic functions in distributed cognitive control. Nature Neuroscience, 20(12), 1669–1679. https://doi.org/10.1038/s41593-017-0020-1
Hardman, A., Töllner, T., & Martinovic, J. (2020). Neural differences between chromatic- and luminance-driven attentional salience in visual search. Journal of Vision, 20(3), 5. https://doi.org/10.1167/jovi.20.3.5
Harrison, B. J., Davey, C. G., Savage, H. S., Jamieson, A. J., Leonards, C. A., Moffat, B. A., Glarin, R. K., & Steward, T. (2022). Dynamic subcortical modulators of human default mode network function. Cerebral Cortex, 32(19), 4345–4355. https://doi.org/10.1093/cercor/bhab487
Herman, J. P., & Krauzlis, R. J. (2017). Color-change detection activity in the primate superior colliculus. Eneuro, 4(2). https://doi.org/10.1523/eneuro.0046-17.2017
Ince, S., Harrison, B. J., Felmingham, K. L., Jamieson, A. J., Davey, C. G., Agathos, J. A., Moffat, B. A., Glarin, R. K., & Steward, T. (2025). Subcortical modulation of the salience network during negative emotional processing in mood and anxiety disorders. Molecular Psychiatry, 30(11), 5475–5485. https://doi.org/10.1038/s41380-025-03135-5
Ince, S., Steward, T., Harrison, B. J., Jamieson, A. J., Davey, C. G., Agathos, J. A., Moffat, B. A., Glarin, R. K., & Felmingham, K. L. (2023). Subcortical contributions to salience network functioning during negative emotional processing. NeuroImage, 270, 119964. https://doi.org/10.1016/j.neuroimage.2023.119964
Jonauskaite, D., & Mohr, C. (2025). Do we feel colours? A systematic review of 128 years of psychological research linking colours and emotions. Psychonomic Bulletin & Review, 32(4), 1457–1486. https://doi.org/10.3758/s13423-024-02615-z
Kanai, R., Komura, Y., Shipp, S., & Friston, K. (2015). Cerebral hierarchies: predictive processing, precision and the pulvinar. Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1668), 20140169. https://doi.org/10.1098/rstb.2014.0169
Keller, G. B., & Mrsic-Flogel, T. D. (2018). Predictive processing: A canonical cortical computation. Neuron, 100(2), 424–435. https://doi.org/10.1016/j.neuron.2018.10.003
Khalaf, A., Lopez, E., Li, J., Horn, A., Edlow, B. L., & Blumenfeld, H. (2025). Shared subcortical arousal systems across sensory modalities during transient modulation of attention. NeuroImage, 312, 121224. https://doi.org/10.1016/j.neuroimage.2025.121224
Kirsch, W., & Kunde, W. (2023). Changes in body perception following virtual object manipulation are accompanied by changes of the internal reference scale. Scientific Reports, 13(1). https://doi.org/10.1038/s41598-023-34311-8
Kok, P., Mostert, P., & de Lange, F. P. (2017). Prior expectations induce prestimulus sensory templates. Proceedings of the National Academy of Sciences, 114(39), 10473–10478. https://doi.org/10.1073/pnas.1705652114
Koole, S. L., & Tschacher, W. (2016). Synchrony in psychotherapy: A review and an integrative framework for the therapeutic alliance. Frontiers in Psychology, 7. https://doi.org/10.3389/fpsyg.2016.00862
Kusztor, A., Mulay, N., Yamada, M., Hohwy, J., & Tsuchiya, N. (2025). Lifting the veil: probing altered visual perception in derealization. Neuroscience of Consciousness, 2025(1). https://doi.org/10.1093/nc/niaf045
Lafer-Sousa, R., Conway, B. R., & Kanwisher, N. G. (2016). Color-Biased Regions of the Ventral Visual Pathway Lie between Face- and Place-Selective Regions in Humans, as in Macaques. The Journal of Neuroscience, 36(5), 1682–1697. https://doi.org/10.1523/jneurosci.3164-15.2016
Lane, R. D., Ryan, L., Nadel, L., & Greenberg, L. (2015). Memory reconsolidation, emotional arousal, and the process of change in psychotherapy: New insights from brain science. Behavioral and Brain Sciences, 38. https://doi.org/10.1017/s0140525x14000041
Lee, C. W., & Cuijpers, P. (2013). A meta-analysis of the contribution of eye movements in processing emotional memories. Journal of Behavior Therapy and Experimental Psychiatry, 44(2), 231–239. https://doi.org/10.1016/j.jbtep.2012.11.001
LeGates, T. A., Altimus, C. M., Wang, H., Lee, H.-K., Yang, S., Zhao, H., Kirkwood, A., Weber, E. T., & Hattar, S. (2012). Aberrant light directly impairs mood and learning through melanopsin-expressing neurons. Nature, 491(7425), 594–598. https://doi.org/10.1038/nature11673
Liu, J., & Zeng, F. (2025). The neural mechanisms of visual and vestibular interaction in self-motion perception. Biology, 14(7), 740. https://doi.org/10.3390/biology14070740
Lynch, E., Dempsey, B., Saleeba, C., Monteiro, E., Turner, A., Burke, P. G. R., Allen, A. M., Dampney, R. A. L., Hildreth, C. M., Cornish, J. L., Goodchild, A. K., & McMullan, S. (2022). Descending pathways from the superior colliculus mediating autonomic and respiratory effects associated with orienting behaviour. The Journal of Physiology, 600(24), 5311–5332. https://doi.org/10.1113/jp283789
McFadyen, J., Mermillod, M., Mattingley, J. B., Halász, V., & Garrido, M. I. (2017). A Rapid Subcortical Amygdala Route for Faces Irrespective of Spatial Frequency and Emotion. The Journal of Neuroscience, 37(14), 3864–3874. https://doi.org/10.1523/jneurosci.3525-16.2017
McKeefry, D. J., & Zeki, S. (1997). The position and topography of the human colour centre as revealed by functional magnetic resonance imaging. Brain, 120(12), 2229–2242. https://doi.org/10.1093/brain/120.12.2229
McPeek, R. M., & Keller, E. L. (2002). Saccade Target Selection in the Superior Colliculus During a Visual Search Task. Journal of Neurophysiology, 88(4), 2019–2034. https://doi.org/10.1152/jn.2002.88.4.2019
Menon, V. (2011). Large-scale brain networks and psychopathology: A unifying triple network model. Trends in Cognitive Sciences, 15(10), 483–506. https://doi.org/10.1016/j.tics.2011.08.003
Menon, V., & Uddin, L. Q. (2010). Saliency, switching, attention and control: A network model of insula function. Brain Structure and Function, 214(5-6), 655–667. https://doi.org/10.1007/s00429-010-0262-0
Murphey, D. K., Yoshor, D., & Beauchamp, M. S. (2008). Perception matches selectivity in the human anterior color center. Current Biology, 18(3), 216–220. https://doi.org/10.1016/j.cub.2008.01.013
Murray, M. M., Lewkowicz, D. J., Amedi, A., & Wallace, M. T. (2016). Multisensory processes: A balancing act across the lifespan. Trends in Neurosciences, 39(8), 567–579. https://doi.org/10.1016/j.tins.2016.05.003
Paulus, M. P., & Stein, M. B. (2010). Interoception in anxiety and depression. Brain Structure and Function, 214(5-6), 451–463. https://doi.org/10.1007/s00429-010-0258-9
Pessoa, L. (2017). A Network Model of the Emotional Brain. Trends in Cognitive Sciences, 21(5), 357–371. https://doi.org/10.1016/j.tics.2017.03.002
Pessoa, L., & Adolphs, R. (2010). Emotion processing and the amygdala: from a ‘low road’ to ‘many roads’ of evaluating biological significance. Nature Reviews Neuroscience, 11(11), 773–782. https://doi.org/10.1038/nrn2920
Phelps, E. A., Ling, S., & Carrasco, M. (2006). Emotion Facilitates Perception and Potentiates the Perceptual Benefits of Attention. Psychological Science, 17(4), 292–299. https://doi.org/10.1111/j.1467-9280.2006.01701.x
Qian, J., Zhang, K., Wang, K., Li, J., & Lei, Q. (2018). Saturation and brightness modulate the effect of depth on visual working memory. Journal of Vision, 18(9), 16. https://doi.org/10.1167/18.9.16
Saalmann, Y. B., Pinsk, M. A., Wang, L., Li, X., & Kastner, S. (2012). The pulvinar regulates information transmission between cortical areas based on attention demands. Science, 337(6095), 753–756. https://doi.org/10.1126/science.1223082
Santos-Mayo, A., & Moratti, S. (2025). How fear conditioning affects the visuocortical processing of context cues in humans. Evidence from steady state visual evoked responses. Cortex, 183, 21–37. https://doi.org/10.1016/j.cortex.2024.11.005
Schmid, M. C., Mrowka, S. W., Turchi, J., Saunders, R. C., Wilke, M., Peters, A. J., Ye, F. Q., & Leopold, D. A. (2010). Blindsight depends on the lateral geniculate nucleus. Nature, 466(7304), 373–377. https://doi.org/10.1038/nature09179
Schneider, L., Dominguez-Vargas, A.-U., Gibson, L., Kagan, I., & Wilke, M. (2020). Eye position signals in the dorsal pulvinar during fixation and goal-directed saccades. Journal of Neurophysiology, 123(1), 367–391. https://doi.org/10.1152/jn.00432.2019
Seeley, W. W., Menon, V., Schatzberg, A. F., Keller, J., Glover, G. H., Kenna, H., Reiss, A. L., & Greicius, M. D. (2007). Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control. The Journal of Neuroscience, 27(9), 2349–2356. https://doi.org/10.1523/jneurosci.5587-06.2007
Sherman, S. M. (2016). Thalamus plays a central role in ongoing cortical functioning. Nature Neuroscience, 19(4), 533–541. https://doi.org/10.1038/nn.4269
Shevell, S. K., & Kingdom, F. A. A. (2008). Color in complex scenes. Annual Review of Psychology, 59(1), 143–166. https://doi.org/10.1146/annurev.psych.59.103006.093619
Sridharan, D., Levitin, D. J., & Menon, V. (2008). A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proceedings of the National Academy of Sciences, 105(34), 12569–12574. https://doi.org/10.1073/pnas.0800005105
Stefanics, G., Heinzle, J., Horváth, A. A., & Stephan, K. E. (2018). Visual mismatch and predictive coding: A computational single-trial ERP study. The Journal of Neuroscience, 38(16), 4020–4030. https://doi.org/10.1523/jneurosci.3365-17.2018
Strumpf, H., Mangun, G. R., Boehler, C. N., Stoppel, C., Schoenfeld, M. A., Heinze, H.-J., & Hopf, J.-M. (2013). The role of the pulvinar in distractor processing and visual search. Human Brain Mapping, 34(5), 1115–1132. https://doi.org/10.1002/hbm.21496
Takakuwa, N., Isa, K., Onoe, H., Takahashi, J., & Isa, T. (2021). Contribution of the pulvinar and lateral geniculate nucleus to the control of visually guided saccades in blindsight monkeys. The Journal of Neuroscience, 41(8), 1755–1768. https://doi.org/10.1523/jneurosci.2293-20.2020
Tamietto, M., & de Gelder, B. (2010). Neural bases of the non-conscious perception of emotional signals. Nature Reviews Neuroscience, 11(10), 697–709. https://doi.org/10.1038/nrn2889
Thayer, J. F., & Lane, R. D. (2000). A model of neurovisceral integration in emotion regulation and dysregulation. Journal of Affective Disorders, 61(3), 201–216. https://doi.org/10.1016/s0165-0327(00)00338-4
van den Heuvel, M. P., & Sporns, O. (2019). A cross-disorder connectome landscape of brain dysconnectivity. Nature Reviews Neuroscience, 20(7), 435–446. https://doi.org/10.1038/s41583-019-0177-6
Vandewalle, G., Collignon, O., Hull, J. T., Daneault, V., Albouy, G., Lepore, F., Phillips, C., Doyon, J., Czeisler, C. A., Dumont, M., Lockley, S. W., & Carrier, J. (2013). Blue light stimulates cognitive brain activity in visually blind individuals. Journal of Cognitive Neuroscience, 25(12), 2072–2085. https://doi.org/10.1162/jocn_a_00450
Vandewalle, G., Schwartz, S., Grandjean, D., Wuillaume, C., Balteau, E., Degueldre, C., Schabus, M., Phillips, C., Luxen, A., Dijk, D. J., & Maquet, P. (2010). Spectral quality of light modulates emotional brain responses in humans. Proceedings of the National Academy of Sciences, 107(45), 19549–19554. https://doi.org/10.1073/pnas.1010180107
Vazquez, S. R. (2005). Peripheral light stimulation for rapid emotional, somatic, and transpersonal regulation. Subtle Energies & Energy Medicine, 16(3), 225–249. https://journals.holosuniversity.org/index.php/seemj/article/view/58/46
Vazquez, S. R. (2009, Winter). A new paradigm for eye movement psychotherapy. Annals of the American Psychotherapy Association, 12(4), 20–29.
Vazquez, S. R. (2014). Emotional transformation therapy: An interactive ecological psychotherapy. Bloomsbury Publishing. https://www.bloomsbury.com/au/emotional-transformation-therapy-9781442238183/
Vazquez, S. R. (2015). Accelerated ecological psychotherapy: ETT applications for sleep disorders, pain, and addiction. Bloomsbury Publishing. https://www.bloomsbury.com/au/accelerated-ecological-psychotherapy-9781442247802/
Wali, M., Lee-Miller, T., Babu, R., & Block, H. J. (2023). Retention of visuo-proprioceptive recalibration in estimating hand position. Scientific Reports, 13(1). https://doi.org/10.1038/s41598-023-33290-0
Wallace, M. T., Woynaroski, T. G., & Stevenson, R. A. (2020). Multisensory integration as a window into orderly and disrupted cognition and communication. Annual Review of Psychology, 71(1), 193–219. https://doi.org/10.1146/annurev-psych-010419-051112
Wampold, B. E. (2015). How important are the common factors in psychotherapy? An update. World Psychiatry, 14(3), 270–277. https://doi.org/10.1002/wps.20238
Wang, Y. C., Bianciardi, M., Chanes, L., & Satpute, A. B. (2020). Ultra high field fMRI of human superior colliculi activity during affective visual processing. Scientific Reports, 10(1). https://doi.org/10.1038/s41598-020-57653-z
Wilms, L., & Oberfeld, D. (2018). Color and emotion: effects of hue, saturation, and brightness. Psychological Research, 82(5), 896–914. https://doi.org/10.1007/s00426-017-0880-8
Zhang, X., Zhaoping, L., Zhou, T., & Fang, F. (2012). Neural activities in V1 create a bottom-up saliency map. Neuron, 73(1), 183–192. https://doi.org/10.1016/j.neuron.2011.10.035
Zhao, Y., Kirschenhofer, T., Harvey, M., & Rainer, G. (2024). Mediodorsal thalamus and ventral pallidum contribute to subcortical regulation of the default mode network. Communications Biology, 7(1). https://doi.org/10.1038/s42003-024-06531-9
Article Information
Published Date
Received: 12 April 2026
Published: 28 May 2026
Cite This Article
Bruno, J. (2026). Sensory-Salience Regulation in Emotional Transformation Therapy: A Clinical-Theoretical Framework. Journal of Psychological Experience. https://doi.org/10.64484/jpsyexp-x4s8m0l2c8n1y8z0
Author(s) Information
Jeffrey Bruno
Visual Ecology and Psychology Research Foundation
2338 Beach Boulevard, Pacifica, California 94044, United States
https://orcid.org/0009-0003-1272-5199
Correspondence: contact@childwisdom.org
Author(s) Contribution
JB: Conceptualization, Investigation, Visualization, Writing - Original Draft, Writing - Review and Editing.
Author(s) Approval
The author declares that they are the sole author of this research and have approved this submission for publication. The author accepts full responsibility for the claims, data, citations, interpretations, and final wording presented here.
Competing and/or Conflicting Interests
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
"The author previously used Emotional Transformation Therapy in clinical practice and currently serves as a board member of the Visual Ecology and Psychology Research Foundation, a nonprofit organization that supports research on visually assisted psychotherapy. The author reports no commercial, proprietary, or financial involvement with ETT training or related entities."
Funding
The author declares that no funding was received for the research, authorship, and/or publication of this article.
Ethics Approval and Consent to Participate
Not applicable – the author declares that this research did not involve human participants and/or animals.
Consent for Publication
Not applicable – the author declares that this research did not include identifiable personal data or images of individuals.
Data Availability
Not applicable – the author declares that no primary data were generated or analysed in this research.
Permission to Use Third-Party Material
The author declares that all figures, tables, and other materials were created by the author(s) and have not been previously published, and that no third-party permissions are required.
Preprint Disclosure
The author declares that this research has not been posted as a preprint.
AI Use Disclosure
The author declares that AI tools were used in the research, writing, or editing of this article.
"AI tools were used in the preparation of this manuscript. ChatGPT (OpenAI) and Claude (Anthropic) were used for language refinement, structural editing, and synthesis support during literature-based theoretical framing. All outputs were reviewed, revised, and verified by the author, who accepts full responsibility for the manuscript’s claims, citations, interpretations, and final wording."
Copyright
© Author(s). Licensee Journal of Psychological Experience. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution, or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. Any usage, dissemination, or reproduction not adhering to these conditions is strictly prohibited.
Publisher's Note
Journal of Psychological Experience (JPsyExp) undertakes all reasonable endeavours to ensure the accuracy of all information (“Content”) contained within its publications. Nevertheless, JPsyExp, its agents (including the editor, members of the editorial team or board, reviewers, and any guest editors), licensors, stakeholders, or third-party vendors, provide no representations or warranties, express or implied, regarding the accuracy, completeness, or suitability of the Content for any particular purpose. Any opinions or views expressed in these publications are those of the authors alone and do not reflect, nor are they endorsed by, JPsyExp. JPsyExp stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. The accuracy of the Content should not be assumed and ought to be independently verified using primary sources. JPsyExp assumes no liability for any loss, action, claim, proceeding, demand, cost, expense, damage, or other liability of any nature, whether arising directly or indirectly in relation to or as a consequence of the use of the Content.